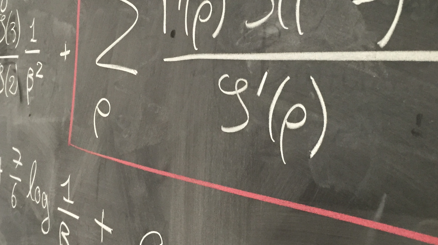Alison Etheridge (Oxford University)
Some mathematical models from population genetics (lecture 3)
Our aim in these lectures is to provide some flavour of the rich variety of mathematical models that arise in the study of the genetics of natural populations and to present some of the main tools that are used to explore their behaviour.
Although the aim is to build a coherent picture of a group of models, each lecture is to a large extent self-contained.
Lecture 1: Although our main focus is spatially distributed populations, we begin with a little history, followed by an introduction to the classical Wright-Fisher and Moran models. This provides the opportunity to introduce some terminology and gain some insight into the nature of the approaches that we’ll take in the more complicated models that lie ahead. Key concepts will be diffusion approximations, genealogical trees, and the notion of duality.
Lecture 2: The next step will be to generalise these models to include spatial structure. This topic has a long and distinguished history, and again we shall begin with classical models due to Kimura, Wright, and Malécot, in which very explicit calculations are possible. However, not only do these models make some very strong assumptions, but in the case of the Wright and Malécot approach to modelling populations evolving in a spatial continuum, in what is arguably the most natural setting of a population evolving in continuus time in a continuous two-dimensional spatial environment, we shall see that there is a mathematical inconsistency, stemming from what is popularly known as ‘the pain in the torus’.
Lecture 3: Our next model, the spatial Lambda Fleming-Viot process (SLFV), is one way to overcome the pain in the torus. Having introduced the SLFV framework, we use it to investigate the way in which spatial structure interacts with a simple form of natural selection. In particular we ask how strong the selection acting on a gene must be relative to neutral mutation if we are to be able to see its trace in genetic data.
Lecture 4: Starting with the work of Fisher (1937), the spread of a selectively favoured gene through a population has been modelled through what is now known as the Fisher-KPP equation. A powerful tool in this context as been duality with a spatial branching process. However, natural selection can take many forms, which leads us to consider more general reaction-diffusion equations than the Fisher-KPP equation, and this leads to a slightly different sort of duality. We illustrate with a model of so-called hybrid zones. We uncover the importance of the shape of the domain in which a population evolves, something which we often ignore in our models.
Lecture 5: In this last lecture we outline some very recent work on a model that attempts to capture some of the local interactions between individuals in the population. Population densities fluctuate in space and time as individuals compete for space and resources. What effect does this have on ancestral lineages, and therefore on the patterns that we see in genetic data?